.jpg)

.png)
.jpg)
.jpg)
.jpg)
- 1
- 2
- 3
- 4
- 5
- 6
- 北京师范大学任海云教授做客细胞发育与未来农业前沿论坛第二十七期 2023-11-24
- 德国马普植物生理所Reimo Zoschke博士做客细胞发育与未来农业前沿论坛第二十... 2023-11-23
- 清华大学刘玉乐教授做客细胞发育与未来农业前沿论坛第二十五期 2023-11-15
- 瑞士日内瓦大学教授Jean-David Rochaix做客植物细胞与发育前言论坛第二十三... 2023-09-24
- 中国科学院遗传与发育生物学研究高彩霞研究员做客植物细胞与发育前沿论坛第... 2023-09-04
- 韩国建国大学尹大珍(Dae-Jin Yun)教授做客细胞发育与未来农业前沿论坛第二... 2023-07-19
- 中国科学院遗传与发育生物学研究所谢旗研究员做客细胞发育与未来农业前沿论... 2023-06-06
- 奥地利科学技术研究院Daniel Zilberman教授做客细胞发育与未来农业前沿论坛... 2023-06-01
- 中国科学技术大学谭树堂教授做客细胞发育与未来农业前沿论坛第十七期 2023-05-28
- 山东农业大学卢从明教授做客细胞发育与未来农业前沿论坛第十六期 2023-05-22
- 荷兰拉德堡德大学须健教授做客细胞发育与未来农业前沿论坛第十五期 2023-05-18
- 中国科学院分子植物科学卓越创新中心刘宏涛研究员做客植物细胞与发育前沿论... 2023-04-04
- Nat Commun-方玉达/涂晓雨团队合作揭示组蛋白H2A通过cohesin靶向进行单泛素... 2023-03-06
- PNAS—薛红卫课题组与合作者揭示磷脂酸细胞器间转运进而调控叶绿体发育的机... 2023-02-02
- Current Opinion in Biotechnology-陆钰明课题组综述植物大规模基因编辑最新... 2023-01-09
-
叶片是植物进行光合作用的主要器官,也是植物生长发育过程中能量和营养物质的主要来源。叶片衰老是植物生长发育的最后一个阶段,有序的叶片衰老对于确保植物繁殖和生存至关重要。叶片衰老是一个受发育和环境因素调控的复杂过程,在高等植物的发育和生命周期中起着关键作用。I型酪蛋白激酶(CK1)是真核生物中高度保守的丝氨酸/苏氨酸蛋白激酶,在细胞增殖、光信号传导和植物激素作用等多种生物学过程中发挥作用。然而,CK1在植物衰老中的生物学功能尚不清楚。
5月4日,中心薛红卫教授团队在New Phytologist发表了题为“Casein kinase 1 AELs promote senescence by enhancing ethylene biosynthesis through phosphorylating WRKY22 transcription factor” 的研究论文,报道了拟南芥I型酪蛋白激酶AELs通过磷酸化转录因子WRKY22促进乙烯生物合成进而促进植物叶片衰老的功能和机制研究。

该团队通过系统的遗传学和生物化学研究,揭示了拟南芥I型酪蛋白激酶AELs介导转录因子WRKY22的磷酸化促进乙烯生物合成进一步调控植物叶片衰老的功能和机制。表型分析显示缺失或过表达酪蛋白激酶AEL的拟南芥分别表现出延迟或加速的叶片衰老表型。进一步的生化证据表明,AELs与WRKY22相互作用并磷酸化WRKY22的Thr57、Thr60和Ser69以增强其转录活性。遗传分析显示WRKY22磷酸化的增加或抑制导致叶片衰老的促进或延迟。ChIP-qPCR和酵母单杂交实验证明WRKY22可以直接与乙烯合成基因ACS7的启动子区结合,促进ACS7基因的转录,并且WRKY22的磷酸化增强了ACS7的转录水平,促进乙烯水平,从而促进叶片衰老。
该研究证明了AEL介导的磷酸化通过增强WRKY22反式激活活性在调节乙烯生物合成和促进叶片衰老中的关键作用,有助于阐明乙烯生物合成的精细调控和完善植物叶片衰老的调控网络。

上海交通大学博士生朱国庆为论文第一作者,薛红卫教授为通讯作者,渠莉副教授参与了工作。该研究得到了国家自然科学基金的资助。
-
青枯菌(劳尔氏细菌,Ralstonia spp.) 通过根部侵染超过54个科250多种植物,造成细菌性枯萎病,也被称为细菌性青枯病。青枯菌寄主范围广泛,包括多种重要的经济作物,如马铃薯、番茄、烟草、茄子、辣椒、花生、香蕉等,在农业生产上危害严重,且难以防治。青枯菌通过根部自然孔口和伤口进入植物维管组织后形成系统性侵染,是典型的维管束病害,对其致病机制一直研究的不够深入。
5月1日,中心余刚副教授课题组和中国科学院分子植物科学卓越创新中心Alberto Macho研究员课题组在Cell Reports发表了题为“Cell wall-mediated root development is targeted by a soil-borne bacterial pathogen to promote infection”的研究论文,揭示了青枯菌分泌三型效应蛋白RipAC靶向纤维素合成复合体相关蛋白CSI1 (cellulose synthase-interactive protein 1),减少纤维素合成,操纵植物侧根发育以增加侵染点的分子机制。

CSI1是植物细胞中连接纤维素合成复合体 (CSC, cellulose synthase complex) 和微管的关键桥梁蛋白,与CSC和微管直接互作,正调控纤维素合成。青枯菌分泌效应蛋白RipAC破坏CSI1与CSC蛋白复合体的形成,负调控纤维素合成。有意思的是,表达RipAC和csi1突变体植物材料均表现出侧根增多的表型。且青枯菌自然侵染情况下,会诱导侧根发生,从而增加侵染点。该研究进一步利用纤维素合成抑制剂isoxaben和侧根诱导化合物BiAux处理植物后,接种青枯菌,均表现出更敏感表型,而植物侧根合成受损突变体均表现为更抗病表型。纤维素合成受影响后,青枯菌能更早更多地诱导侧根发生,表明了纤维素合成与侧根发生的相关性,及其对青枯菌侵染的影响。
综上所述,该研究揭示了土传病害青枯菌利用效应蛋白操纵植物侧根发育,从而增加侵染点而致病的分子机制,增加了对于土传病害操纵植物寄主发育的理解。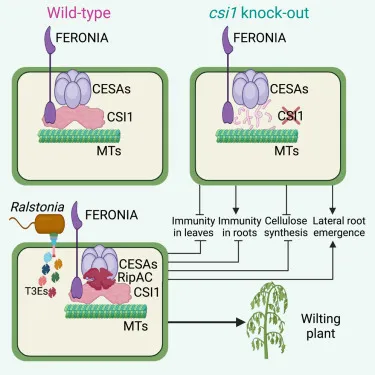
青枯菌操纵植物侧根发育分子机制模型
该项工作由余刚课题组和中国科学院分子植物科学卓越创新中心Alberto Macho课题组合作完成,余刚为本文第一作者和共同通讯作者,Alberto Macho研究员为本文共同通讯作者。中国科学院分子植物科学卓越创新中心博士生张露,硕士生薛好、陈玉娇,分子植物科学卓越创新中心Rosa Lozano-Duran研究员 (现为德国图宾根大学教授),赵春钊研究员和刘鑫博士后,西班牙马德里理工大学Juan C. del Pozo研究员都参与了该工作。本研究得到国家自然科学基金,中科院先导计划等基金资助。
-
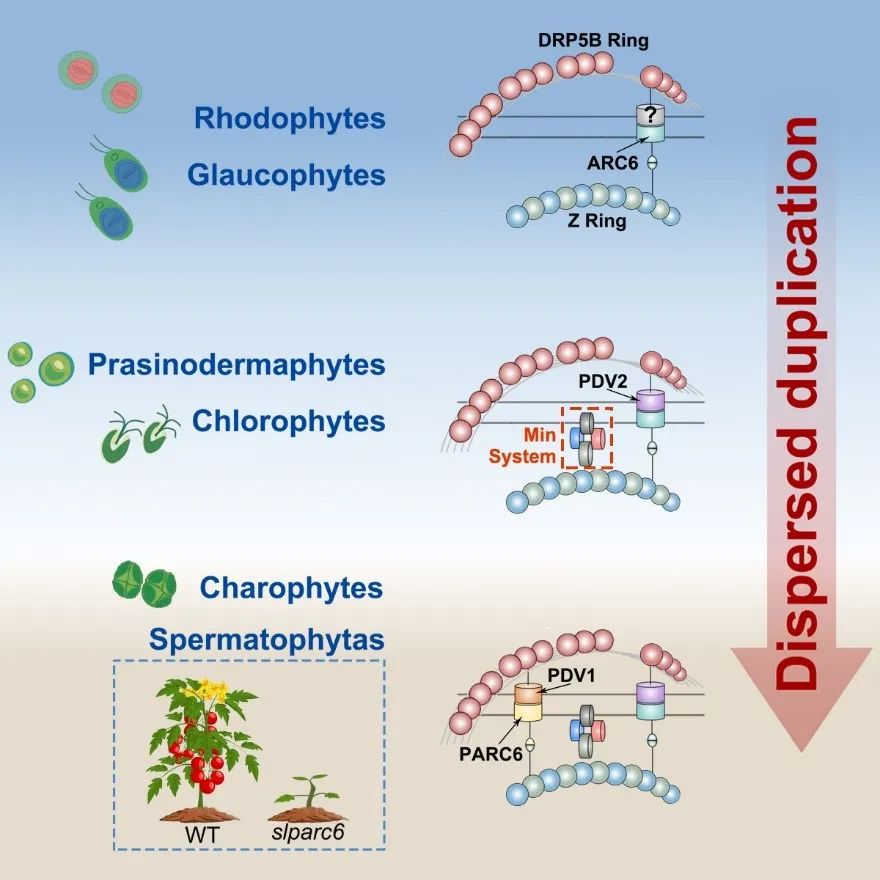
近日,上海交通大学农业与生物学院/上海交通大学单细胞生物学联合研究中心陈诚课题组在Cell Reports发表了题为“Adaptive Evolution of Chloroplast Division Mechanisms during Plant Terrestrialization”的研究论文,揭示了植物叶绿体分裂机制中少数起源于真核宿主细胞的分裂基因更多地参与到帮助植物适应登陆后复杂、多变环境生物学过程的适应性演化机制。
叶绿体起源于蓝细菌与真核宿主细胞的内共生作用,是绿色植物特有的重要细胞器,在光合作用等基本生命活动中发挥关键作用。在漫长演化历程中,植物逐渐形成了调控叶绿体数量和大小的功能机制,以确保细胞内环境的稳定,并在植物生长发育过程中提供其所需光合作用能力。尽管叶绿体分裂已经得到广泛研究,但其核心成员的起源和功能演化仍不清楚。
本研究利用最新公布的泛植物界物种的基因组和转录组数据,鉴定了不同物种中存在的叶绿体分裂核心成员,并重建了它们的演化史。将完整分裂环结构首次出现的时间点从以往仅限于维管植物,推前到水生植物轮藻中。区别于大多数功能机制的演化,叶绿体分裂是在植物登陆前(例如灰藻)就受到了环境的强烈选择,基因的分散复制为此提供了适应性演化的主要动力。通过利用多组学数据(转录组、蛋白质组、转录调控、单细胞等)开发的基因功能注释工具,进一步表明分散复制演化的叶绿体分裂基因获得了更多物种特异性功能。结合遗传和生理实验,证明了由分散复制演化的叶绿体分裂重要成员PARC6在番茄中获得了除叶绿体分裂功能之外调控植物生长发育的新功能。
综上,本研究为叶绿体分裂机制的演化提供了一个综合视角,并强调了基因的分散复制作为叶绿体分裂适应性演化主要来源的潜力。
上海交通大学农业与生物学院/上海交通大学单细胞生物学联合研究中心刘默洋副研究员和余晶硕士(已毕业)为论文第一作者,陈诚(长聘教轨)副教授为通讯作者,博士生杨鸣和上海交通大学生命科学技术学院曹凌雁助理研究员参与了研究工作。该研究得到国家自然科学基金、上海市浦江人才计划等项目的资助。
论文链接:https://doi.org/10.1016/j.celrep.2024.113950
-
线粒体基因的表达、生物生成和功能实现主要依赖于核基因编码蛋白。这一过程主要通过转录后水平进行调控,包含了广泛的内含子剪接事件。在高等植物中,绝大多数线粒体内含子属于能够自我剪接的II类内含子。然而,由于植物线粒体基因的异质性,其内含子失去了自我剪接的能力,因而必须依靠核编码的蛋白质因子来辅助完成剪接过程。这些剪接因子参与内含子剪接的特异性高度可变,一些剪接因子(即特定剪接因子)参与一个或少数几个内含子剪接;另一些剪接因子(即通用剪接因子)参与了数量众多的内含子剪接事件。然而,这些剪接因子如何参与剪接反应的分子机制尚不清楚。
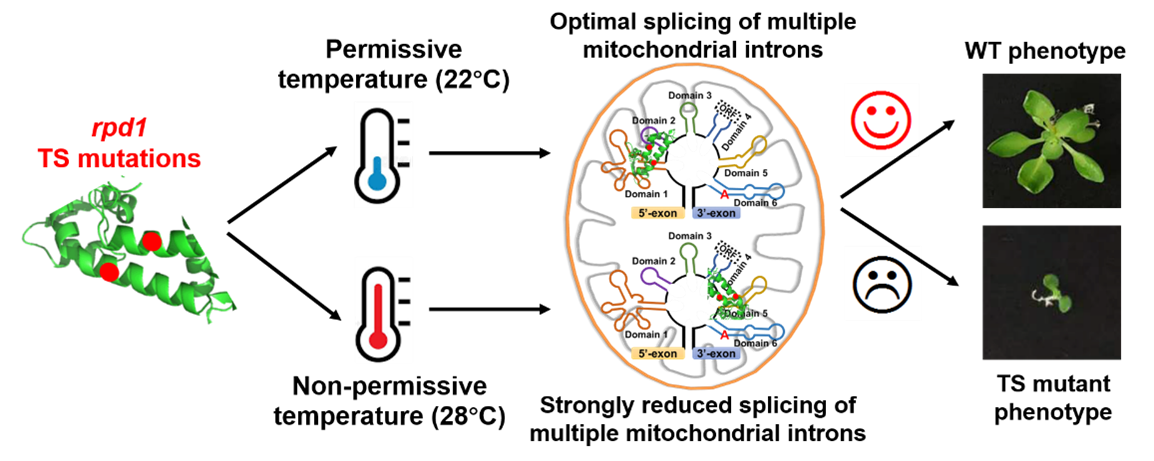
2月14日,上海交通大学单细胞生物学联合研究中心王传德课题组在核酸领域顶级期刊 Nucleic Acids Research 发表了题为 “Temperature-sensitive splicing defects in Arabidopsis mitochondria caused by mutations in the ROOT PRIMORDIUM DEFECTIVE 1 gene”的研究论文,揭示了PORR蛋白家族的RPD1蛋白直接参与多个线粒体内含子的剪接。研究指出,RPD1基因的两个温度敏感性突变对其参与内含子剪接的功能产生了影响,从而证明了高温条件下突变体的不定根发育异常主要是由于RPD1与线粒体内含子的结合能力降低,进而损害了线粒体的功能,而并非由于RPD1直接参与调控拟南芥不定根的形成。此外,RPD1蛋白与目标内含子的多个不同区域相结合,表明RPD1在内含子剪接过程中可能具有多样性机制。

RPD1基因最初因其在调控植物根原基发育中的作用而被发现,其编码蛋白对根系发育具有关键影响。因突变导致根发育异常,故命名为Root Primordium Defective 1。该基因的两种温度敏感突变体在常温(22°C)下能形成不定根,而在高温(28°C)下则不能。进一步研究发现,RPD1蛋白是一个包含15个成员的蛋白家族中的一员,该家族中有三个成员随后被发现参与了线粒体或叶绿体内含子剪接,引发了RPD1可能参与细胞器RNA转录后调控的猜想,尽管其在根发育中的确切作用仍然不明。
为阐明RPD1的分子功能,首先确定了RPD1蛋白在细胞中的定位,发现其主要定位于线粒体,这与之前的核定位假设相悖。对rpd1部分回补突变体植株的深入分析发现,RPD1对于多个线粒体的内含子剪接至关重要,从而证实了其在线粒体剪接中的广泛作用。这一作用与RPD1 的敲除突变体所展现的呼吸链缺陷及胚胎致死现象相一致。此外,在不同温度(22°C和28°C)下对rpd1温度敏感突变体的分析显示,高温(28°C)加剧了线粒体内含子剪接的缺陷,显著降低了线粒体活性,进而抑制了包括根原基细胞在内的细胞分裂。根发育的抑制主要由线粒体功能障碍引起,而非RPD1基因直接参与根形态构建。通过RIP-seq分析确认了RPD1直接与线粒体内含子相结合,且其结合位点跨越不同内含子的多个区域,而这些结合序列间并无显著同源性,这暗示了RPD1参与内含子剪接过程的复杂性与多样性。
上海交通大学王传德教授为论文的第一作者,法国国家农业科学院Hakim Mireau资深研究员为通讯作者。
该研究是王传德团队在细胞器基因表达调控领域的又一重要进展。近期发现核编码蛋白参与细胞器内含子剪切 (PNAS, 2020;Nucleic Acids Research, 2024);较系统地阐明了PPR蛋白参与线粒体RNA 的3’端加工成熟的多样性分子机制 (Nucleic Acids Research, 2017; Plant Physiology, 2022; Nucleic Acids Research, 2023);揭示了油菜核编码蛋白Rfo在翻译水平上抑制不育基因的表达,从而恢复Ogura-CMS的育性 (PNAS, 2021)。






 loading......
loading......